

任何細胞執行其功能,以及存活下去都需要能量代謝,T細胞自然也不例外。
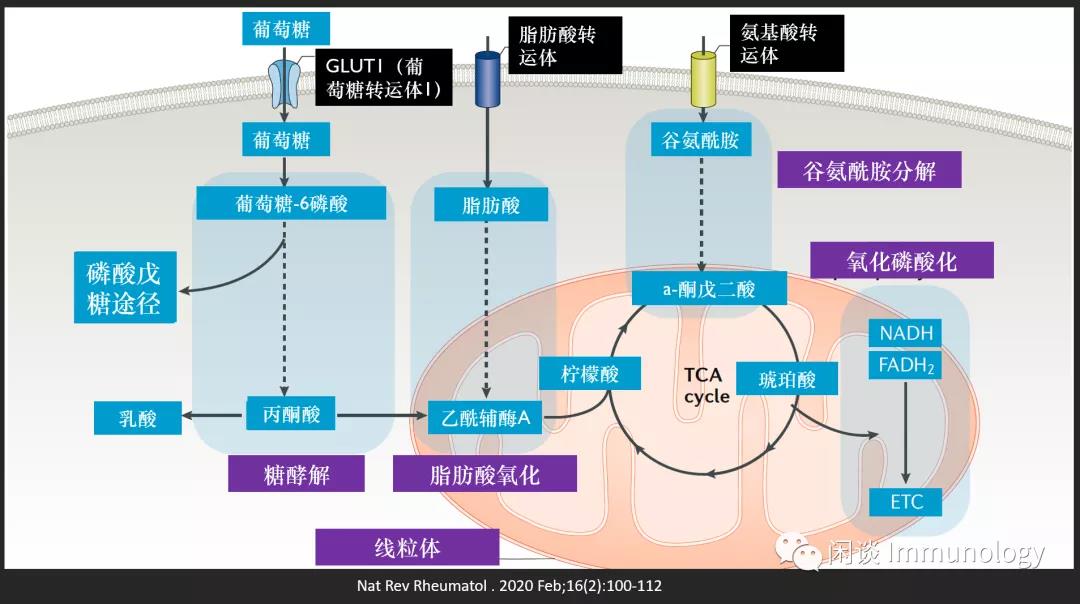
葡萄糖參與糖酵解和氧化磷酸化。糖酵解發生在細胞質中,將葡萄糖轉化為丙酮酸(產生兩個分子 ATP),繼而轉化為乳酸或乙酰輔酶A (在線粒體中)。 脂肪酸代謝也會產生乙酰輔酶A。
乙酰輔酶A隨后進入三羧酸(TCA)循環,并通過氧化磷酸化產生ATP。
除了葡萄糖代謝, 免疫細胞可以利用TCA循環機制,通過代謝谷氨酰胺來產生能量等。除了產生能量,各種中間分子還是合成脂質,核酸和非必需氨基酸的前體。
TCR結合之后,T細胞啟動合成代謝,從靜息狀態進入激活,增殖。線粒體生物合成,提供能量供效應T細胞。此過程中PI3k-AKT-mTORC活化, 誘導T細胞代謝重編程,從脂肪酸氧化和丙酮酸氧化轉為糖酵解和谷氨酰胺分解。 并產生中間產物:核苷酸, 氨基酸和脂肪酸 。
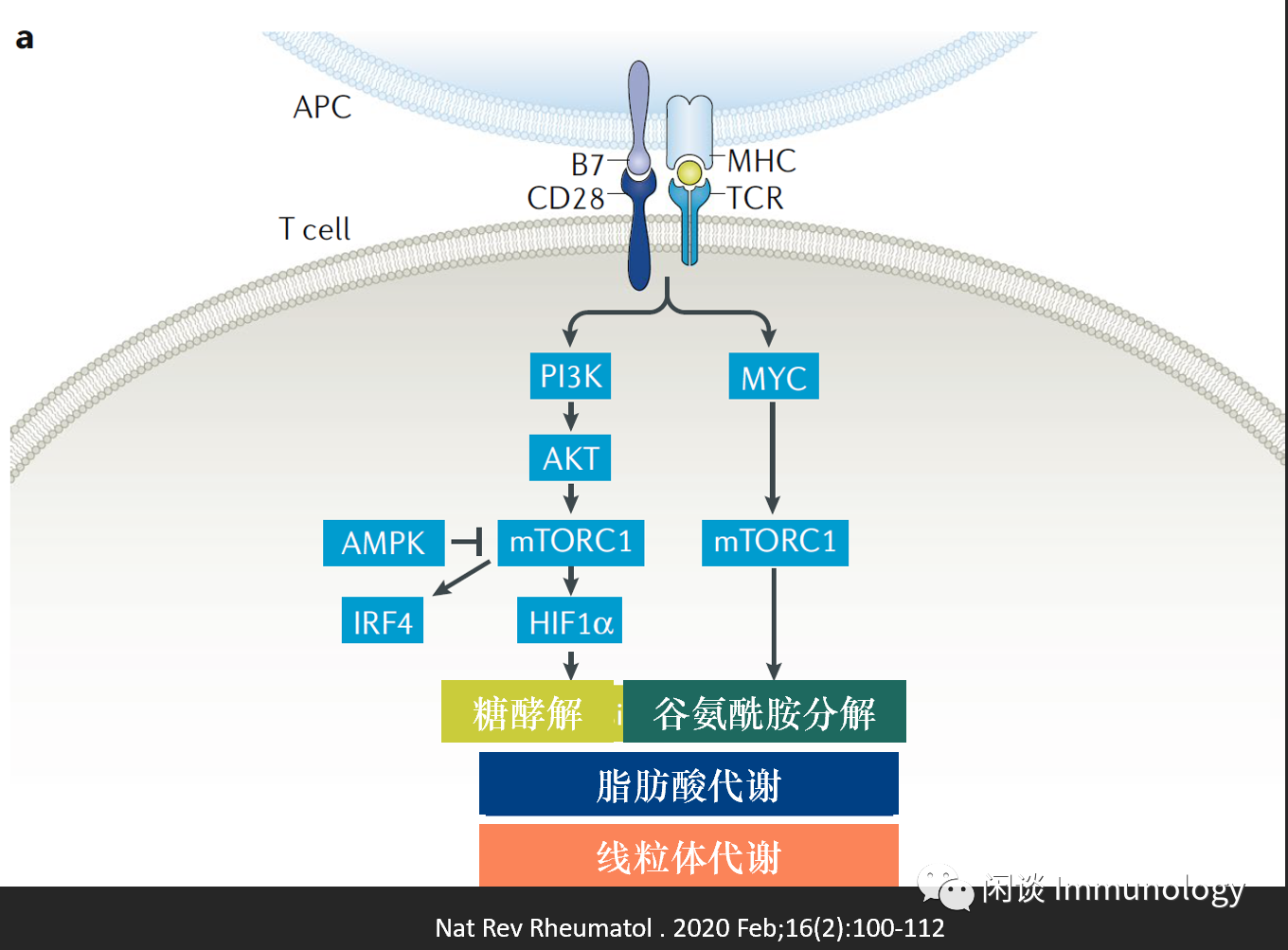
T細胞活化后的分化,受到MYC(通過mTORC調控代謝)的調控。高表達MYC的細胞分化為效應T細胞,低表達MYC的細胞分化為記憶樣T細胞。
糖酵解是T細胞活化后所需的主要代謝通路(Warburg effect:從氧化磷酸化轉化為糖酵解)。T細胞增殖需要線粒體代謝,產生ATP給生物合成,信號通路,產生ROS,激活NFAT(IL-2產生的關鍵轉錄因子)等。
naive CD4+T 細胞激活后,分化為效應T細胞亞群: TH1,TH2,TH17,TFH等,他們根據功能需要使用不同的代謝途徑。mTORC依然是最主要的調節通路。
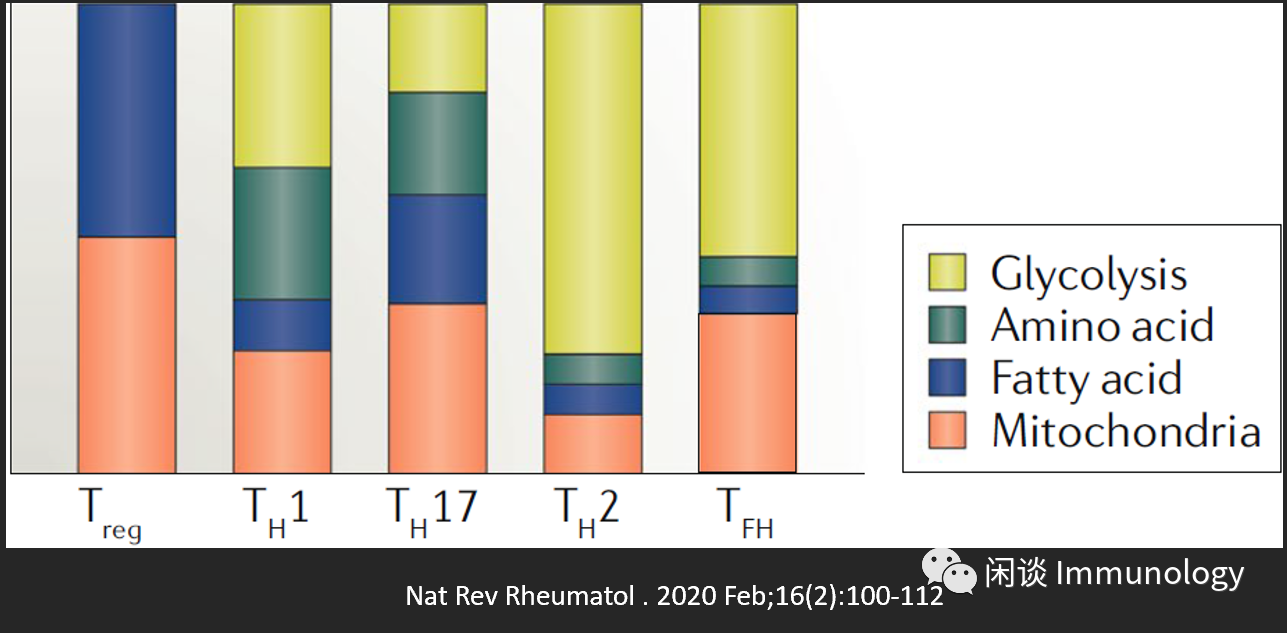
mTORC1活性對于TH1,TH17, CD8+T分化極其重要。
mTORC1-mTORC2活性則對TH2和TFH重要。
一些T細胞分化重要的調節分子,也被證實參與了代謝通路,如:ICOS(
inducible T cell co-stimulator);GLUT1( glucose transporter 1);IRF4(interferon regulatory factor 4 ),BCL6。
效應T細胞需要糖酵解,GLUT1在其中有重要作用。
效應T細胞的命運受到葡萄糖代謝調控分子(HIF1α和PDH)的調節。
丙酮酸生成后(糖酵解的最后一步),丙酮酸通過乳酸脫氫酶(LDH)轉化為乳酸,通過PDH轉化乙酰輔酶A進入線粒體。 HIF1α調節CD4和CD8T細胞的糖代謝,同時也控制TH1和TH17細胞的分化;相反,mTORC2調節RhoA(小GTPase),是TH2分化必需的。
TH17表達ICER(inducible cAMP early repressor),上調PDK1,從而產生高水平乳酸(低水平乙酰輔酶A)。
氨基酸對于蛋白質合成,核酸合成,調節mTORC1通路,T細胞應激通路都是必需的。
谷氨酰胺是一種非必需氨基酸,是循環中最豐富的氨基酸,在活化的T細胞中消耗增加。
谷氨酰胺分解途徑中,谷氨酰胺被水解成谷氨酸,它被進一步代謝為α酮戊二酸(三羧酸循環的中間體,組蛋白去甲基化酶和DNA去甲基化酶的底物),促進向效應T細胞分化。谷氨酸缺乏,抑制α酮戊二酸產生,則促進向調節T細胞分化。 谷氨酰胺酶是谷氨酰胺分解途徑第一個酶,被ICER誘?